The Evolution of Early Reptiles
During the Carboniferous (360-285 mya), the amphibians dominated most
terrestrial landscapes. In Carboniferous times, the continents were coming together
to form a more-or-less continuous land mass. Lush forests of seed ferns and
gymnosperms grew in lowland swamps in the northern continents. These forests
were populated by giant millipedes (some up to 6.5 feet -or 2 m- long), dragonflies
the size of sea gulls, and an assortments of giant cockroaches, spiders and scorpions.
The southern continent (Gondwanaland) was too cold to support much life and an
ice sheet covered its southern tip (like Antarctica today).
However, small lizard-sized vertebrates, called cotylosaurs, were also in
existence, creeping in and out of vegetation in drier areas, in search of small insects
and worms. They laid eggs that did not have to hatch in water. These were the first
reptiles and they included the ancestors to the other reptiles, birds and mammals.
I. Reptilian Grade of Evolution
The features that distinguish reptiles from amphibians are:
A. The Amniotic Egg
Among the vertebrates, one of the most important events was the
achievement of reproduction independent of water. This was accomplished by the
evolution of a shelled egg and internal extraembryonic membranes.
These have left no trace in the fossil record, but their presence in all groups
of reptiles and their descendants (mammals and birds) strongly supports the
assumption they evolved in a common ancestor.
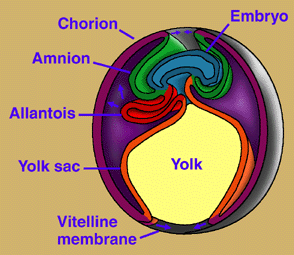
Amniotic eggs are different from the gel coated eggs of amphibians in that
they have:
1. semipermeable shells which allows gases to pass in (oxygen) or out (carbon
dioxide), but keeps fluid in to protect the embryo from desiccation.
2. specialized membranes that lie outside the embryo: the chorion, amnion, and
allantois.
a. the chorion surrounds the embryo and the yolk sac (contains food and
fluid for the embryo.
b. the amnion surrounds the embryo (hence the name amniotes)
c. the allantois grows out of the embryo's lower digestive track and rapidly
covers the surface of the chorion. The interface between the two membranes
is well supplied with blood vessels and acts as a respiratory organ that allows
the diffusion of gases. Nitrogenous wastes of the embryo are deposited into
the allantoic cavity as relatively insoluble, nontoxic precipitates such as uric
acid that the animal need not immediately eliminate.
As the embryo develops, the proteinaceous food in the yolk sac dwindles and
the allantois fills up.
B. Shortened digits, more powerful ankle and wrist bones. More motile head -
reptiles have one occipital condyle on the back of the skull, whereas amphibians
have two.
C. Heart: Amphibians have a single ventricle and two auricles, whereas the
reptilian ventricle is at least partially divided:

This keeps oxygenated and unoxygenated blood from mixing.
D. Thick scaly skin that is keratinized and impermeable to water. Keratin is a
protein found in hair, fingernails and feathers. The protective skin prevents water
loss but is shed periodically as it dries out and the cells die.
E. Excretion: Nitrogenous waste in reptiles is uric acid which can be concentrated
(therefore the animals do not need large amounts of water to get rid of
nitrogenous waste). Remember - In amphibians, urine is quite dilute and may
contain much ammonia.
F. In many amphibians, a single excretory duct system services both the gonads
and the kidneys, whereas reptiles have separate ducts for each system.
G. Stretch receptors within the axial and appendicular skeleton.
H. Expandable rib cage helps pump air into the lungs.
I. Recall that temperature fluctuates much more on land than in water. The first
reptile were like amphibians in that they caould not regulate their body
temperature - they are cold blooded (ectothermic). They use the sun to warm up
and shade to cool down.
The First Reptiles
The earliest reptiles, the cotylosaurs, are from the early Carboniferous (350
mya) of Scotland,and includes the genera Hylonomus and Paleothyris from the
mid-carboniferous (310 and 300 mya respectively) from Nova Scotia.

These remains in Nova Scotia were discovered in the 1850s and have yielded 100s of
fossil vertebrates since. It seems that in mid Carboniferous times this area of Nova
Scotia was covered with lush forests of vascular plants up to 30 m tall. Occasionally
the lakes flooded and the forests were inundated with sediment. The trees died and
fell, leaving only their roots and buried lower trunks in place. As new forests
became established above, the centers of the tree trunks rotted, and arthropods,
molluscs (snails), amphibians, and small reptiles fell in. These lived for some time
in the tree trunk and the amphibians and reptiles fed on the invertebrates (as
evidenced by insect parts in the fecal remains of the tetrapods), but eventually they
died and were preserved as fossils inside the protective trunk.
The body of these early reptiles is slender and about 200mm long (including
the tail). Compared to many amphibians, the head is relatively small (1/5 the body
length as apposed to 1/3 to 1/4). But the jaws are stronger - new sets of muscles
appear that are capable of pulling the jaw up and out. The jaw construction and
teeth suggest that they fed on invertebrates, probably insects, millipedes and
centipedes. They were agile insectivores, rather like modern lizards in their ecology.
Carboniferous Reptiles
There are Four major groups of amniotes (tetrapods with an amnion), three
of which had evolved by the late Carboniferous.
These animals can be distinguished on the basis of the number and position of extra
openings in the skull.
Why would these openings appear? --
The argument is that bone is costly to produce (it takes a lot of calcium and
phosphate) and it is heavy, therefore it can be advantageous to dispense with it
where it is not needed. Much of the skull is under stress from the movements of
the jaws and neck muscles, but some spots, in the cheek region, are under no stress,
and cavities (or fenestrae) can develop without reducing the effectiveness of the
skull.
The groupings are:
- 1. Anapsida - amniotes with no temporal fenestrae.
- Example - Cotylosaurs - early forms like Hylonomus and Paleothries,
and the turtles.
- 2. Synapsida - amniotes with one temporal fenestrae (surrounded by the
postorbital, jugal, and squamosal bones)
- Example - mammals
- 3. Diapsida - amniotes with two temporal fenestrae (one as in synapsids, and
the other surrounded by the postorbital, squamosal, and parietal).
- Example - Lizards, snakes, birds, crocodiles and many extinct forms
including dinosaurs.
- 4. Euryapsid - amniotes with one temporal fenestrae (as in second one in
diapsids - thought to be through secondary loss - evolved later than
carboniferous)
- Example - many extinct marine animals plesiosaurs and icthyosaurs.

Permian Amniotes
During the Permian, the continents moved into close contact to form a single large
supercontinent, Pangea. The polar ice cap melted and the lands that had made up
the southern Gondwanaland continent moved northward and became warmer.
In the Permian, the southern continent was occupied by both amphibians and
reptiles. The land that had been the northern continents, on the other hand,
became hot and arid in many parts. The humid forests of ferns were replaced by
seed-bearing conifers. During this time, the amphibians were replaced by the
reptiles as the dominate land vertebrate. This change must, in part, be due to the
climatic changes to dryer conditions.
I. Synapsids
The dominate early reptiles were the synapsids. They were the most important
group of fully terrestrial tetrapods in the Permian.
1. Pelycosaurs
The most diverse tetrapods of the early Permian, representing up to 70% of all
genera, were the pelycosaurs.
Many of these had very elongated (up to 3 meters) neural spines on the
cervical and dorsal vertebrae which supported a large sail on the back. A well
known example is the genus Dimetrodon.

The function of this sail is a fascinating topic for paleobiological speculation.
During growth, the neural spines of something like Dimetrodon became relatively
more and more elongate in proportion to the increase in body weight of the animal,
but not in proportion to body length. We have already seen in other animals, that
increase in size results in volume (or weight) increases as a cube of the body length,
whereas surface area increases as a square of the body length. The fact that the sail
increases in length in proportion to body volume indicates it has a physiological
function related to offsetting problems of large body weight - such as heat or water
balance.
Does the construction of the sail give us any clues as to the function of the
sail?
- 1. The neural spines have grooves at the base that were probably occupied by
blood vessels.
- 2. The neural spines always lie in a neat fence-like array indicating they were
held together by a tough covering of skin.
These two clues, and the probability that the pelycosaurs were cold-blooded
indicates that the sail was used to warm the body quickly by basking in the sun.
Smaller cold blooded animals can regulate their body temperature with
behavioral modifications which would be inefficient in a larger animal. One
scientist calculated that it would take a 250 k Dimetrodon 12 hours of basking in
the sun to warm its body temperature from 25° to 30°C, but only 3 hours with a
sail.
As a top predator, these few extra hours may have permitted Dimetrodon to grab
its still-torpid prey which lacked sails. Furthermore, higher activity levels give
them greater hunting efficiency, greater food intake, faster digestion, and faster
growth.
Locomotion: There was no well-defined ankle joint, and the toes were long and
splayed out sideways as the animal walked. The feet thus had no forward thrust
but instead simply supported the limbs on the ground. The forelimbs were
passive supports that prevented the animal from falling on its face, while the
hindlimbs provided all the thrust in walking from powerful muscles that rotated
the femur at the hip joint.
The backbone of the pelycosaurs was stiff and did not bend from side to
side as much as in the amphibians or the cotylosaurs. This is an important
advance in solving what is called the Carrier Constraint (named after David
Carrier of the U of Michigan who described it). David Carrier pointed out that
the sprawling locomotion of a salamander or even a lizard (although its legs are
more under the body than a salamander) that depend on side to side movement
of the spinal column to propel the legs forward (this is inherited from the fish
ancestors where the movement of the tail provides forward thrust) compresses
each lung alternatively as it moves. This means they can't run and breath at the
same time (they run then stop and breathe).

Pelycosaurs solved this problem by stiffening the vertebral column and allowing
less side to side movement. Locomotion comes from rotating the legs to the
forward position - not throwing them forward using the backbone.
Feeding: Despite their clumsy gait, early pelycosaurs were all carnivores. The
jaws were simply hinged and could slam shut on a prey - there was no side-to
side chewing possible. But this structure does make it possible to take a hold of
prey. Prey were probably killed outright by the large dagger-like teeth when the
jaw slammed shut. They had massive jaw muscles and may have been able to
hold struggling prey in their mouths until they died.
During the late Permian, some pelycosaurs were vegetarians. These
animals were the first terrestrial vegetarian vertebrates. The teeth in the jaw of
these pelycosaurs are blunt and there were rows of palatal teeth on the roof of the
mouth. The jaw hinge is worn down and the jaw could move forward and
backward. Plant material could thus be ground between the palatal teeth and the
teeth in the jaw or the tongue. Plant material is difficult to digest and low in
protein. Animals that eat plant material must consume large quantities to get
enough nutrients or to selectively feed on the most nutritious parts of the plants
- nectar, fruit, and seeds. In the Permian, the dominate plants were the
gymnosperms (e.g., conifers) which do not have fruits or flowers (hence no
nectar). Herbivorous reptiles had to consume a lot of plant material and as a
consequence large body sizes began to appear. (Large carnivores switch to plants
because they have a big enough gut to survive - e.g., bears to pandas).
One lineage of the pelycosaurs gave rise in the mid-Permian to another major
synapsid group - the Therapsida. All the pelycosaurs were extinct by the end of
the Permian.
2. Therapsida
These varied in size from small lizard-like animals to large rhinoceros-sized
animals. Understanding therapsid evolution is difficult because they evolved
and differentiated so rapidly in the Permian. They may have been so successful
and evolved so rapidly because of modifications to the jaws and teeth allowed
them to exploit new food sources and to compete well for others.
The number of bones in the jaw and the way it is hinged with the skull changed.
In most other amniotes, the lower jaw is made up of several small bones,
in the therapsids, this reduced to 2 large bones and 3 smaller ones.
In typical reptiles, the lower jaw hinges on the skull in the very back: it
forms a simple flap which can only go up or down. In the therapsids, the hinge
is placed forward and it allows more complex movements of the jaw (chewing,
not just biting and swallowing). They also had differentiated teeth that included
large canines.
The skull is often modified with horns, thickened plates and cranial crests.
Indicates a lot of behavioral interaction such as fighting, ritual defense or
competition for territory or mates. etc.
Therapsids had a stiffer spine and longer legs than the pelycosaurs - this gave
them more efficient locomotion.
Therapsids evolved first in the temperate zones of the world and were especially
successful in what are now the southern continents. The Therapsids obviously
experimented with temperature control and may have been the first warm
blooded (homeothermic) animals. What is our evidence? Fossils indicate the
presence of fur (modified scales) and a layer of fat under the skin.
II. Diapsids
The synapsids dominated the Permian, but other groups were present too.
The diapsids began to radiate and a number of new forms appeared. Many lineages
existed in the Permian and went extinct at its end without leaving any descendants.
Although many of these are fascinating in their biology, we are going to focus on
just two that had a major impact on future vertebrate evolution.
1. Archosaurs - In the Permian these medium sized carnivores fed on fish and
amphibians and smaller synapsids. They all have another opening on their skull
behind the nostril.
2. Early Lepidosaurs - Ancestors to the lizards, snakes and tuatara (True lizards
appeared in the mid-mesozoic, the first tuatara appears in the Triassic). They lived
much like the living lizards but some were fantastic. One group were the gliding
weigeltisaurs of Europe and Madagascar. These small reptiles have enormously
elongated ribs which stick out sideways but could be folded back when the animal
was running about. The ribs were presumably covered with skin and they could
have glided from tree to tree much as a flying squirrel does today.
The Mass extinction at the End of the Permian
The biggest mass extinction of all time took place at the end of the Permian
(Maxwell 1989). Of the 37 tetrapod families present in the last 5 years of the
permian, 27 went extinct (a loss of 75%). Only 10 families survived!
This loss of tetrapod families must count as a mass extinction since so many groups
died out, and across such a broad ecological spectrum, from small carnivores to
massive herbivores. At the same time, 50% of marine invertebrate families died
out, the highest rate of extinction in the history of the seas.
The fossil record is not clear so we do not know if the extinction was sudden and
catastrophic or more gradual with a loss of 5-6 families every million years.
Hypotheses:
- 1. creation of Pangea cut down on amount of coastline and continentalized
the climate.
- 2. Asteroid or comet shower (little evidence but it can't be ruled out).