


The ciliates (phylum Ciliophora) is one of the largest groups of protists and contains those forms with the most complex cell structures.
Most protists are microscopic, ranging in size from about 5 to 500µ. A few, such as the foraminiferans, are much larger and commonly visible to the naked eye. Some protists are larger than the smallest of the animals and they include marine, freshwater, terrestrial, and symbiotic (including pathogenic) species.
Protists are not all closely related to each other. Some are actually plants and others animals or fungi, while some are just ... Well... Just their own separate groups. They were traditionally put together into a kingdom before electron microscopy and molecular biology revealed their true diversity.
Antony van Leeuwenhoek is generally credited with being the first person to report seeing protists in about 1675. In fact, Leeuwenhoek was the first to describe a number of microscopic aquatic life forms (protozoa, rotifers, and others), referring to them as "animalcules" ("little animals"). The name protozoon (Greek, proto, "first"; zoon, "animal") was coined by Goldfuss in 1818 and included protists, but also sponges, cnidarians, rotifers, and some small worms.
Following the discovery of cells in 1839, the distinctive nature of protists became apparent. On the basis of this distinction, von Siebold, in 1845, restricted the name Protozoa to apply to all unicellular forms of animal life. In the late 1940s, algae, protozoa, and some other groups were united into a single group, the Protista by TL Jahn, which is today recognized in many introductory biology books as a kingdoms of organisms. However, it is now apparent that it is not monophyletic but is instead a paraphyletic group. In other words, the kingdom Protista is itself not united by any unique distinguishing features, or synapomorphies. Protists are definable only as a eukaryotes lacking multicellular of organization seen in plants, animals and fungi (shared primitive feature).
Throughout most of the twentieth century, a relatively standard classification scheme developed for the protists. This scheme was based on the idea that the different groups could be classified primarily by their modes of nutrition and locomotion. Thus, they were divided into:
These are the divisions that you learned in introductory biology. While these divisions might accurately describe protists' role in ecosystems, they do not accurately reflect evolutionary relatedness.
Pseudopodia and flagella are present in many different kinds of cells (including plant and animal cells) and their presence does not indicate unique relatedness (i.e., they are shared primitive features or symplesiomorphies).
Photosynthetic protists contain many different types of chlorophylls and have differently constructed chloroplasts, indicating that they are not closely related.
Although there continues to be much heated debate over how these enigmatic organisms are related to each other - for example one of the most exciting recent discoveries, for example, was that the former protistan phylum Myxozoa actually comprises a group of highly modified cnidarians that are parasites of certain invertebrates and vertebrates. This revelation was made possible by the discovery of certain metazoan and cnidarian features (e.g., collagen, nematocysts) in these animals, as well as ribosomal DNA data.
Most experts agree that there are several well-defined groups of protists which are summarized in the next section. You are not responsible for these groups for the exam - this information is just for your interest:
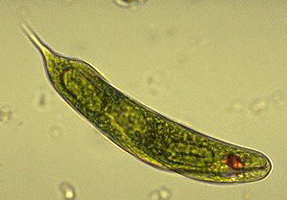

The euglenids (e.g., Euglena) include many photosynthetic species and so were once thought to be related to other phytoflagellates. Studies of the cells with electron microscopy, has shown that they have a unique chloroplast structure that is not similar to that found in any other photosynthetic protist. Instead, the flagellum has a lattice-like supporting rod and the cell membrane is underlain by ribbons of unique microtubules -- two synapomorphies that link them to the Kinetoplastids. Kinetoplastids (e.g., Trypanosoma and other parasites) are heterotrophic flagellates and were once thought to be related to other zooflagellates. Kinetoplastids get their name from their unique mitochondrion - it is a single, large branching mitochondrion with a large amount of DNA that is visible under the light microscope (this DNA consists of small circles linked together to form a relatively huge mass).
Alveolata - Three protists groups (ciliates, apicomplexans, and dinoflagellates) have an
alveolar membrane system, which consists of flattened membrane-bound sacs (alveoli) lying
just underneath to the usual outer cell membrane. This, together with recent evidence from
molecular sequence comparisons, indicate that these three protist phyla are evolutionarily
closely related.
The ciliates (phylum Ciliophora) is one of the largest groups of protists and contains those
forms with the most complex cell structures.
Phylum Apicomplexa includes many important parasites (e.g., Plasmodium - the causative
agent of malaria) -- they have a complex structure at the apical end that is used to penetrate
the host cells (hence their name).

Stramenopiles all have flagella with fine hairs protruding from it. This group includes some
photosynthetic forms (e.g., diatoms, brown algae, golden algae - all with chlorophyll a and c2)
as well as some saprophytic forms that were once considered to be unicellular fungi.

Rhizopoda or amebas with broad pseudopodia.



Diplomonads are bi-flagellated heterotrophic protists that lack mitochondria and golgi.
They are thought by some to be relatively primitive, but may lack these features because
they live in anaerobic environments (e.g., Giardia).
Chlorophyta or green algae (e.g., Chlamydomonas, Volvox) are actually plants even though they are unicellular.
Microsporidia are all small intracellular parasites of other eukaryotes (including other protists). They contain a unique coiled structure that is extruded from the cell and allows them to penetrate their host's cells. They are of growing medical importance - many are now found in immuno-compromised people. Their so highly modified for living in other cells, that they lack any features that can clearly tell us who they are related to.
Choanoflagellata are filter feeding heterotrophic flagellates that are closely related to sponges. Some workers actually put them in the animal kingdom.
Small Size: Even in the largest multicellular organisms, the regulation of exchanges across cell membranes and the metabolic reactions along the surfaces of various cell organelles are the phenomena on which all life ultimately depends.
Consequently, the total area of these important surfaces must be great enough relative to the volume of the organism to provide adequate exchange and reaction sites.
Without an efficient mechanism of circulation within the body, or the presence of membrane partitions (multicellularity) to enhance and regulate exchanges of materials, protists must remain relatively small; the diffusion distance between their cell membranes (their "body surface") and the innermost parts of their bodies can never be so great that it prevents adequate movement of materials from one place to another within the cell or for the movement of gases and waste in and out of the cell body. Certainly there are structural elements (e.g., microtubules, endoplasmic reticula) and various processes (e.g., protoplasmic streaming, active transport) that supplement passive phenomena. But the fact is, unicellularity mandates that a high surface area:volume ratio be maintained by restricting shape or size. This is the principle behind the fact that the largest protists have assumed shapes that are elongate, thin, or flattened - that maintain small diffusion distances.
Osmotic Regulation: The elimination of metabolic wastes and excess water, especially in
freshwater forms living in hypotonic environments, is facilitated by contractile vacuoles.
These consist of membrane-bound tubes than empty into a large collecting vacuole.
Microtubules suround the tubes and the vacuole - these tubules contract around the tubes
pushing their contents into the vacuole. Then the microtubules around the vacuole contract
and force the water out through a pore on the cell surface.

Nutrition: Various types of nutrition occur among protists. Indeed, they may be either autotrophic or heterotrophic (some may be both). Photosynthetic protists have chloroplasts and are capable of photosynthesis, although not all use the same chloroplast pigments and they often have different chloroplast structure.
Heterotrophic protists require organic chemicals from their environment. Heterotrophic forms may be saprobic, taking in dissolved organics by diffusion, active transport, or pinocytosis. Or they may be holozoic, taking in solid foods such as organic detritus or whole prey by phagocytosis.
All heterotrophic protists acquire food through some interaction between the cell surface and the environment. Those that engage in pinocytosis or that engulf solid food rely on the formation of membrane-bounded vesicles called food vacuole. These structures may form at nearly any site on the cell surface, as they do in the amebas, or at particular sites associated with some sort of "cell mouth," or cytostome, as they do in most protists with more-or-less fixed shapes. The cytostome may be associated with further elaborations of the cell surface that form permanent invaginations or feeding structures.
Once a food vacuole has been formed and has moved into the cytoplasm, it begins to
swell as various enzymes and other chemicals are secreted into it. The vacuole first becomes
acidic, and the vacuolar membrane develops numerous inwardly directed microvilli. As
digestion proceeds, the vacuolar fluid becomes increasingly alkaline.

Sensation: Many protists display remarkable degrees of sensitivity to environmental stimuli and are capable of some fairly complex behaviors. Response behavior may be a function of the general sensitivity and conductivity of protoplasm, or it may involve special organelles. Cilia and flagella are touch- and chemical- sensitive organelles; when mechanically stimulated, they typically stop beating or beat in a pattern that moves the organism away from the point of stimulus.
Most protists react positively or negatively to various chemicals or concentrations of chemicals. For example, amebas are able to distinguish food from nonfood items and quickly egest the latter from their vacuoles. Many ciliates, especially predators, have specialized patches of sensory cilia that aid in finding prey, and even filter feeders use cilia around the cytostome to "taste" and then accept or reject items as food.
Photosynthetic protists typically show a positive taxis to low or moderate light intensities, an obviously advantageous response for these creatures. They usually become negatively phototactic in very strong light. Specialized light-sensitive organelles are known among many flagellates, especially the photosynthetic ones. These eye spots or stigmata (Sing. stigma) are frequently located at or near the anterior end. Some, however, are found associated with the chloroplasts. Eye spots vary in complexity, ranging from very simple pigment spots to complex, lens-like structures.
Reproduction: A major aspect of protistan success is their surprising range of reproductive strategies. Most protists have been able to capitalize on the advantages of both asexual (cell division) and sexual reproduction (exchange of genetic material).
In summary, protist diversity and success reflect the tremendous potential for
variation within the unicellular body. Many of these features are inherited and remain in
multicellular eukaryotes. As we discuss the evolution of animals and plants, you will see that
many of the first animals and plants to evolve retain the cell functions found in protists. But
as time passes, these functions are taken over increasingly more complex organ systems.