- 2. 800 mya trace fossils of burrowing organism from North Australia.
- 3. Not recognizable as extant taxa.
Therefore our theories come from living taxa.
All agree metazoan came from protozoa because of similarity of cell structure. But
beyond this there is little agreement. There are basically two theories for the origin of
animals:
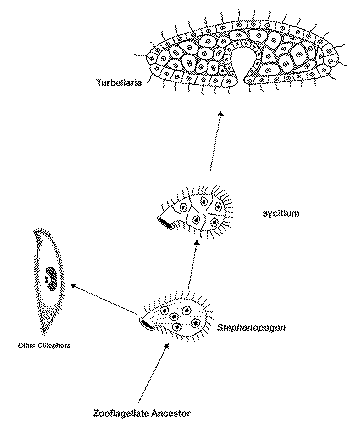
Syncitial Theory for the Origin of Animals
This theory makes three points:
2. The flatworms (like planaria) arose by cellularization of the cytoplasm of a large, multinucleate ciliated protist
3. Sponges arose separately from flagellates.
Supposed homologies:
Stephanopogon is not related to animals or to ciliates and therefore can not be the living representative of the ancestor to either group.
According to this theory the animals:
1. Are monophyletic (sponges and cndarians are related to the rest of the animals).
All these organisms share:
Within this theory are two subtheories:
1. Plakula theory
2. Blastae theory
1. Plakula Theory
This theory was originally proposed by Butschlei (in 1884) when he stated that he believed that the most primitive metazoan after the sponges would be a flattened, disk-shaped, benthic organism.

In recent times this story has been supported by Karl Grell through the discovery of
Trichoplax.

Cross-section through a placozoan:

This story is supported by many marine ecologists who believe that nutrient distribution requires primitive metazoans to have been benthic.
Problem:
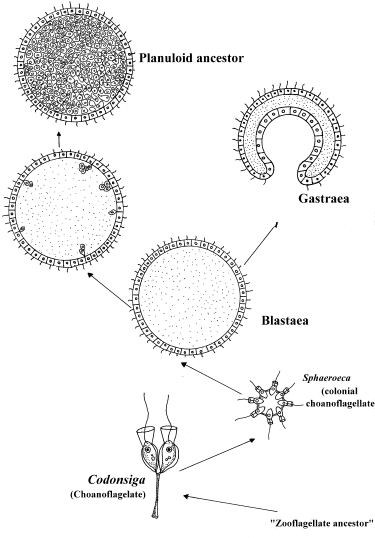
2. Blastae Theory
Haeckel (1874) - starting with the principle that ontogeny recapitulates phylogeny,
there must be stages comparable to:
One of the main differences between unicellular and multicellular organisms is cell differentiation. That is, cells become specialized for a specific function and, usually, take on a characteristic morphology. For example, your body has epithelial cells lining most of its surfaces. If you take an epithelial cell and grow them in tissue culture they will multiply but they will remain epithelial cells.
How do cells get locked into one type? In the early 1900's, scientists suggested that the genes that control a particular kind of cell (such as an epithelial cell, for example) are passed on to those cells during development and all other genes (such as those that control muscle cells) are filtered out. As you well know, that isn't the case - each of your cells has all the genes needed to make an exact copy of you. This means that some genes are not used in some cells -- for example, muscle cell genes are not used in epithelial cells.
Cell differentiation then depends on different genes being active in different cells. This occurs through a process called gene regulation. In gene regulation, one gene (called a regulator gene) acts a a switch that turns other genes on or off. A basic version of this process occurs in bacteria and protists as well as multicellular organisms.
Locking a cell into one type
In some protists (ciliates have been particularly well studied), some genes get turned off and stay off even after mitosis. The whole genome doesn't get reactivated until the protist goes through meiosis. This occurs when either a protein or a methyl group is added to the DNA blocking transcription of the gene at that site. When the DNA is replicated in mitosis, an enzyme copies the methyl group onto the new piece of DNA. When the cell goes through meiosis, however, the methyl groups are removed and the entire DNA is reactivated.
Master Control Genes of Differentiation
In the first steps of development in multicellular organisms, cells divide and
spread but there is no differentiation. Embryos at this stage do, however, have an
anterior to posterior gradient that appears to be inherited from the egg's cytoplasm.
This gives the embryo an anterior "head" end and a posterior "tail" end.
Cell differentiation is under the control of a set of master genes known as the
Hox genes. The Hox genes are a family of genes arranged in a sequence on a
chromosome. The anterior-most gene controls the head end of the body by
regulating which genes in the anterior cells are activated and which are methelyated
(and so turned off). Developmental biologists have investigated the role of these
genes in embryos of fruit flies by turning hox genes on and off - for example, by turning
off the first gene and turning on the second gene in the anterior end of the animal they
can cause legs instead of antenae to grow out of the head.
All animals have Hox genes, but animals that evolve later have more genes
and more complex, segmented bodies. Also the genes that are controlled change
over time - it may be that the origin of new phyla occurs because of these switches.